PROGETTO PREISTORIA
Capitoli da un manuale inedito di storia per le scuole superiori
2.2. Il cespuglio dell’evoluzione umana
«Luce sarà fatta»: le prime ricerche paleoantropologiche
La CAPACITÀ CRANICA misura il volume all’interno del cranio, grosso modo corrispondente alle effettive dimensioni del cervello.
Nell’Origine delle specie, la cui prima edizione viene data alle stampe nel 1859, Darwin glissa prudentemente sulla questione evolutiva dell’origine dell’umanità al fine di evitare polemiche e si limita a notare che «luce sarà fatta sull’origine dell’uomo e sulla sua storia». Due anni prima erano stati scoperti in Germania i primissimi resti fossili capaci di generare dibattiti animati sul passato dell’umanità. L’interpretazione di questi fossili, attribuiti nel 1864 alla nuova specie Homo neanderthalensis (dal nome della località dove furono rinvenuti i fossili, ossia la valle di Neander, in tedesco “Neandertal”), viene però sviata dal fatto che i reperti appartengono a un individuo affetto da vari problemi di salute. Nonostante l’elevata capacità cranica* del reperto, si impone quindi l’immagine degli uomini delle caverne come di claudicanti e scimmieschi bruti preistorici.

I Neanderthal, da esseri bestiali a esseri umani. A DESTRA, una delle primissime illustrazioni di Neanderthal, risalente al 1909; a SINISTRA, una ricostruzione a distanza di un secolo (2010) ispirata dalla recente scoperta di piume e artigli di corvidi e di rapaci probabilmente utilizzati a scopi ornamentali dalle popolazioni eurasiatiche di Neanderthal. FONTI: (a sinistra) particolare da un’opera originale di Fran Kupka (1909), ripresa da Erik Trinkaus e Pat Shipman (1993). “Neandertals: Images of Ourselves”. The Antioch Review 51(3): 194-201. https://doi.org/10.2307/4612796; illustr. a p. 197; (a destra) Mauro Cutrona, “il volto nuovo dell’uomo di Neandertal: Le penne come simbolo di potere” (Grotta di Fumane, provincia di Verona), da Archeoveneto.
Nel libro L’origine dell’uomo e la selezione sessuale, pubblicato nel 1871, Darwin torna sulla questione e suggerisce che le antropomorfe africane debbano essere considerate le parenti prossime di H. sapiens. Inizialmente però ha maggior successo una spiegazione alternativa, secondo cui la culla dell’umanità sarebbe piuttosto da individuarsi in Asia.
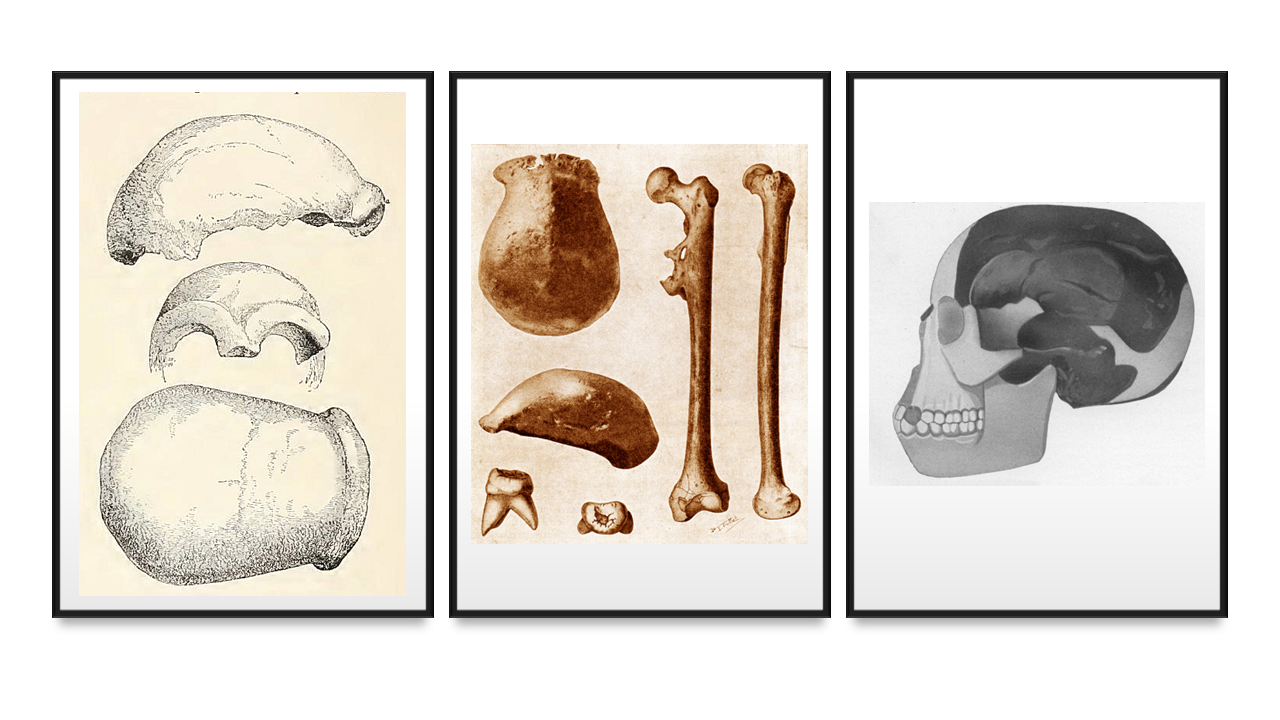
I primi fossili umani e la prima frode paleoantropologica. A SINISTRA: a calotta cranica del primo esemplare di Neanderthal dalla Germania; AL CENTRO: i tre fossili originari provenienti da Giava e descritti da Eugène Dubois (una calotta cranica, un molare e un femore); A DESTRA: ricostruzione del cranio del cosiddetto Eoanthropus o uomo di Piltdown, una frode perpetrata ai danni della comunità scientifica internazionale. FONTI: Neanderthal, Wikipedia; fossili da Giava, Wikipedia; uomo di Piltdwon, Wikipedia (da J. Arthur Thomson, The Outline of Science, 1922).
Pare avvalorare questa ipotesi il ritrovamento di alcuni fossili (poi rivelatisi non particolarmente arcaici) sull’isola indonesiana di Giava, e soprattutto la scoperta del cosiddetto Uomo di Piltdown, in Inghilterra: il reperto consta di un cranio umano e di una mandibola dai tratti chiaramente scimmieschi, il che sembra confermare agli addetti ai lavori sia l’idea dell’Eurasia come culla dell’umanità sia l’ipotesi che lo sviluppo del cervello dovette precedere e guidare gli altri cambiamenti tipici di H. sapiens.

Cranio dell’esemplare giovanile di Australopithecus africanus descritto da Raymond Dart nel 1925 (a SINISTRA); ricostruzione (a DESTRA). FONTI: cranio – Didier Descouens [CC 4.0], da Wikipedia; ricostruzione – © Emiliano Troco.
Fino agli anni Trenta del XX secolo molti paleoantropologi, che avevano scartato a priori il suggerimento darwiniano riguardo alla priorità evolutiva dell’Africa per via di pregiudizi razziali e di sciovinismo imperialistico, continuarono ad andare in cerca degli antenati dell’umanità in Asia. Anche quando nel 1924 fu scoperto in Sudafrica l’eccezionale cranio di un Australopithecus africanus di appena quattro anni di età, non si volle cedere sull’origine eurasiatica di H. sapiens: il cranio venne erroneamente catalogato come il fossile di una semplice antropomorfa dal cervello piccolo.
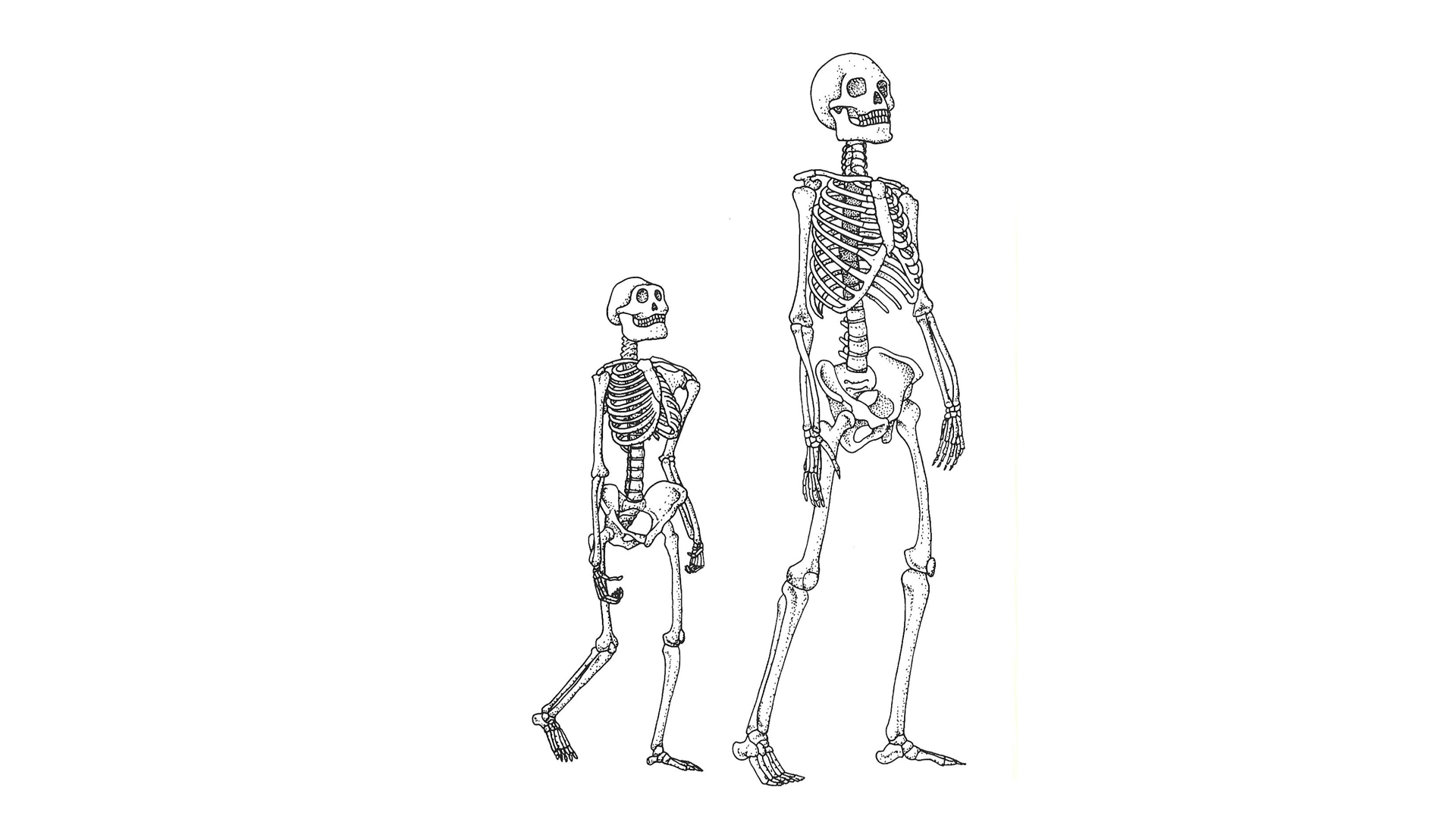
Struttura scheletrica di uno degli esemplari meglio conosciuti di Australopithecus afarensis (a SINISTRA) e di un Homo sapiens moderno (a DESTRA). FONTE: C. Stringer e P. Andrews (2006). Storia completa dell’evoluzione umana. Modena: Logos, p. 188.
Intanto vengono alla luce diversi reperti africani di qualità eccezionale, tra i quali un nuovo cranio appartenente a un esemplare adulto di A. africanus dotato di una capacità cranica superiore a quella di uno scimpanzé (485 cm3 contro circa 300 cm3). Eppure, la comunità scientifica resta scettica fino all’inizio degli anni Cinquanta, quando le nuove tecniche di datazione radiometrica permettono di rivelare che il reperto dell’Uomo di Piltdown non è altro che una frode realizzata unendo furbescamente una calotta cranica umana moderna con una mandibola di orango.

Il paleoantropologo etiope Zeresenay Alemseged mostra un reperto fossile appartenente a un giovane Australopithecus afarensis risalente a 3,3 milioni di anni e scoperto presso Didika, in Etiopia, nel 2000. FONTE: M. Benton (a cura di) (2008). The Seventy Great Mysteries of the World. Londra: Thames & Hudson, p. 53. © Kenneth Garrett.
Cade così l’idea che l’aumento delle dimensioni del cervello sia stato il motore primo dell’evoluzione umana. Nel frattempo, anche i più facinorosi avversari dell’ipotesi africana ammettono di essersi sbagliati. Da allora, il numero dei generi africani scoperti e descritti è aumentato esponenzialmente, al punto che oggi siamo in grado di ricostruire con una buona approssimazione le tappe principali dell’evoluzione degli omìnini umani.
Dalla foresta alla savana: le australopitecine
Esistono molti modi di essere “umani”. L’evoluzione umana non è una marcia inesorabile da forme “primitive” e incomplete a forme “migliori” e più recenti.

Il cespuglio dell’evoluzione umana. FONTE: traduzione e adattamento di Leonardo Ambasciano, diagramma originale di T. Michael Keesey [CC 2.0], da Flickr.
La NICCHIA ECOLOGICA è la posizione occupata da un organismo all’interno del suo ambiente, alla costruzione della quale esso stesso contribuisce attivamente tramite comportamenti e abitudini.
Quello a cui assistiamo è invece il dispiegarsi di un cespuglio di forme differenti e spesso coabitanti nelle medesime zone geografiche, ciascuna adattatasi alla propria nicchia ecologica* attraverso originali sperimentazioni adattative e tramite un miscuglio di caratteri antropoidi e tratti più spiccatamente umani.

Il folto cespuglio dell’evoluzione degli omìnini umani aggiornato al 2019. Nuovi fossili e nuove scoperte paleogenetiche permettono di aggiornare costantemente le relazioni filogenetiche tra queste specie. LEGENDA dei generi elencati: S.: Sahelanthropus; O.: Orrorin; Ar.: Ardipithecus; Au.: Australopithecus; P.: Paranthropus; K.: Kenyanthropus; H.: Homo. Nota bene: a seconda dei criteri considerati, alcune specie sono talvolta considerate sinonimi di altre: H. georgicus è considerato da alcuni paleoantropologi H. erectus; H. ergaster include i primissimi rappresentanti africani di H. erectus; H. rudolfensis è inglobato in H. habilis; il genere Kenyanthropus è considerato talvolta uno dei primissimi rappresentanti di Homo assieme a H. rudolfensis; H. rhodesiensis è ritenuto sinonimo di H. heidelbergensis; la datazione di H. naledi è incerta. FONTE: modificato da Bernard Wood (2019). Human Evolution: A Very Short Introduction. 2nd Ed. Oxford: Oxford University Press, p. 83. Una versione meno aggiornata dello schema è stata pubblicata nella prima edizione disponibile in italiano come Evoluzione umana, Codice edizioni, Torino 2008, p. 82.

Cranio di Sahelanthropus. FONTE: Didier Descouens [CC 4.0], da Wikipedia.
Sahelanthropus dalla Repubblica del Ciad e Orrorin dal Kenya, risalenti rispettivamente a 7 e 6 milioni di anni, possiedono alcune caratteristiche condivise con gli altri omìnini (riduzione dei canini, bipedismo facoltativo) e sono entrambi prossimi al progenitore comune di pànini e omìnini umani.
Ardipithecus dall’Etiopia risale a una forbice temporale compresa tra 5,8 e 4,4 milioni di anni fa e testimonia sia un marcato adattamento fisiologico per una più efficiente locomozione bipede sia una capacità cranica compresa tra i parametri degli attuali pànini (300-350 cm3). La causa dietro alla pressione selettiva nei confronti di forme alternative di bipedismo più o meno facoltativo è probabilmente da ricercarsi nel cambiamento ecologico che segna l’Africa orientale in questo stesso periodo, quando le foreste lasciano spazio agli spazi aperti tipici della savana punteggiati da macchie di vegetazione e di alberi.
Tuttavia, questi tre generi sono noti attraverso fossili frammentari e pertanto la loro posizione filogenetica è ancora soggetta a discussioni specialistiche. Con il genere Australopithecus, invece, i dati a disposizione diventano numerosi e molto più affidabili. Le varie specie assegnate a questo genere hanno una capacità cranica di circa 400-500 cm3 e occupano con successo una fascia territoriale che copre tutta l’Africa orientale fino al Sudafrica tra 4,2 e 1,4 milioni di anni fa.

Evoluzione tecnologica. La produzione di pietre scheggiate prodotte dagli omìnini come utensili. LEGENDA: MAF: milioni di anni fa. FONTE: traduzione e adattamento di Leonardo Ambasciano; schema originale di T. Michael Keesey da Kane, J., E. Willoughby & T. Michael Keesey (2016). God’s Word or Human Reason? An Inside Perspective on Creationism. Portland: Inkwater Press, p. 205; unifacciale del Lomekwiano ripreso da Harmand, S., Lewis, J., Feibel, C. et al. (2015). “3.3-million-year-old Stone Tools from Lomekwi 3, West Turkana, Kenya”. Nature 521: 310-315, p. 313. https://doi.org/10.1038/nature14464.
L’ICNOLOGIA studia le tracce lasciate dagli esseri viventi come conseguenza dei loro comportamenti, tra cui impronte, feci e tane.
Tra i resti attribuiti alle australopitecine abbiamo pietre scheggiate e utilizzate come strumenti da taglio e un eccezionale reperto icnologico*, ossia una serie di impronte fossili risalenti a 3,66 milioni di anni fa e impresse su fine polvere di origine vulcanica presso Laetoli in Tanzania. Questo reperto conferma una locomozione pienamente bipede.

Nuove impronte di australopitecine descritte nel 2016 e rinvenute presso Laetoli, in Tanzania. FONTE: Masao, F. I. et al. (2016). “New Footprints from Laetoli (Tanzania) Provide Evidence for Marked Body Size Variation in Early Hominins”. eLife Sciences 5. https://doi.org/10.7554/eLife.19568.
Homo erectus: fuoco, dieta e viaggi intercontinentali
Mentre una linea di australopitecine evolve in forme più massicce e adattate alla masticazione di cibi più coriacei come radici e tuberi (forme assegnate al genere Paranthropus), un’altra linea di australopitecine più gracili sfuma nei primissimi rappresentanti attribuibili al genere Homo. Entriamo in un territorio tassonomicamente complicato dove le differenze diventano così sottili e le continuità filogenetiche così fluide da rendere discutibile la demarcazione in generi differenti. H. habilis e H. rudolfensis sono per certi versi così simili anatomicamente alle australopitecine – con le quali dovettero condividere spazi e luoghi – che alcuni studiosi pensano debbano rientrare in quest’ultimo gruppo. Diffuse tra 2,4 e 1,5 milioni di anni fa, queste specie hanno una capacità cranica di circa 600-700 cm3. Si tratta di numeri significativi, tanto che alcuni paleoantropologi riconoscono in questo periodo l’esito di un primissimo cambiamento che lega l’aumento delle dimensioni del cervello a mutate abitudini alimentari, e in particolare al consumo di carne e di grassi animali.

Selezione rappresentativa del cranio di alcuni ominidi (non in scala); la freccia del tempo va da sinistra (specie meno recenti) verso destra (specie più recenti). FONTE: Composizione di Leonardo Ambasciano. Fotografie da Phillip L. Walker e Edward H. Hagen, Human Evolution: A Multimedia Guide to the Fossil Record. Student Edition CD-ROM. W.W. Norton & Co © 2003 Regents of the University of California.
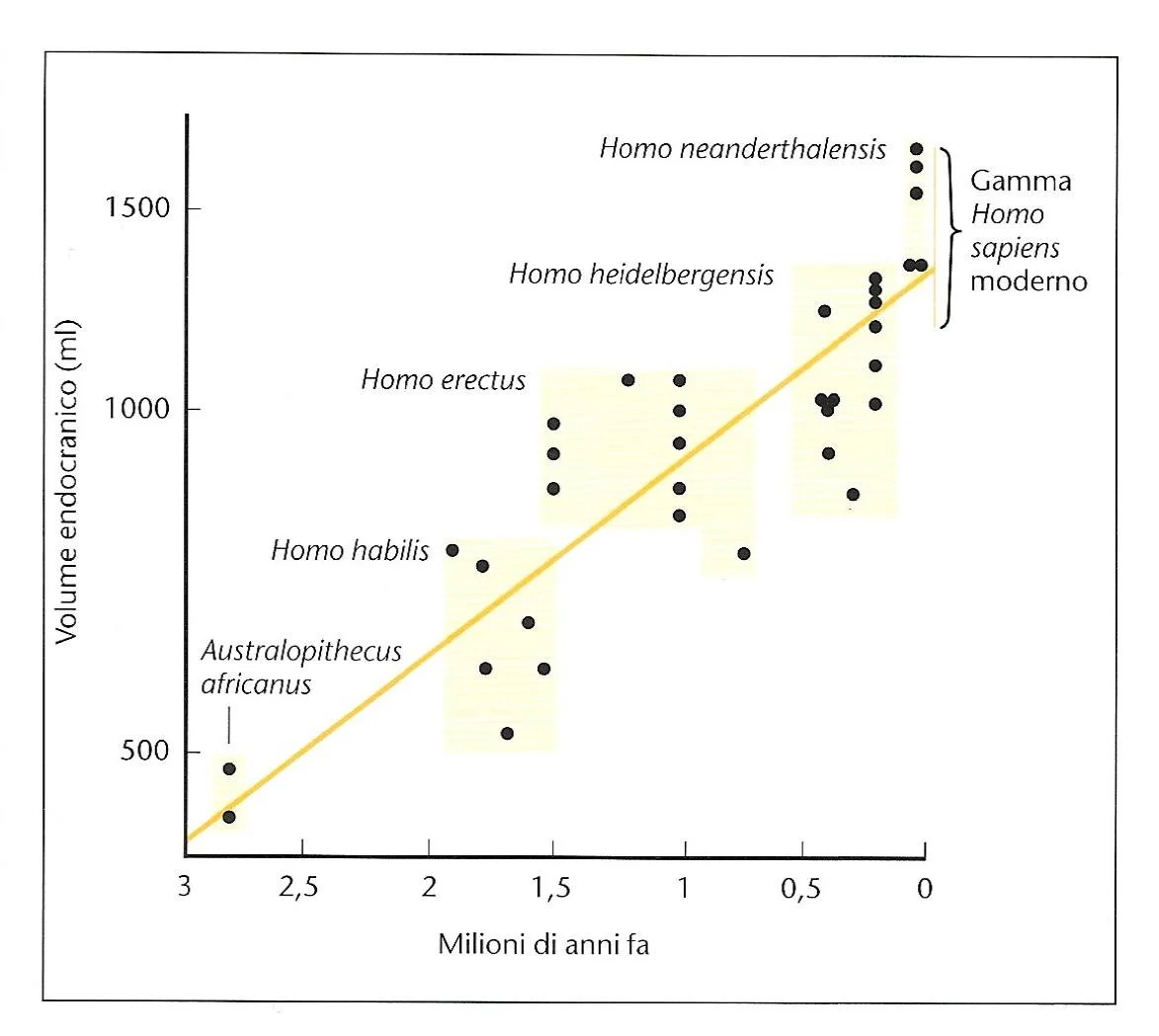
Aumento della capacità cranica degli ominidi nel corso degli ultimi 3 milioni di anni. Ogni specie mostra una gamma di variazioni significative al suo interno. L’aumento del volume del cervello è correlato alla necessità di tenere conto mentalmente dei complessi rapporti sociali all’interno di gruppi sempre più numerosi. FONTE: C. Stringer e P. Andrews (2006). Storia completa dell’evoluzione umana. Modena: Logos, p. 131.
Un secondo cambiamento di rilievo ha luogo con Homo erectus. Si tratta del primo genere di ominine umano per il quale disponiamo di dati relativi alla produzione del fuoco. Il cervello è un organo piccolo (corrispondente al 2% della massa corporea) ma estremamente costoso in termini di consumo metabolico, poiché esige circa il 20% dell’energia necessaria per il funzionamento del corpo. L’uso del fuoco per cuocere gli alimenti comporta benefici significativi – oltre a ridurre la pericolosa carica batterica degli alimenti, questa operazione si traduce nel guadagno minimo di circa il 30% di apporto energetico aggiuntivo in termini di nutrienti e calorie. Al consumo di cibo cotto fa da contraltare la riduzione graduale di denti e delle dimensioni dell’apparato digerente (liberando così energia per il cervello), assieme all’attivazione di enzimi digestivi e di particolari geni preposti alle attività metaboliche. Questa modificazione radicale della nicchia ecologico-alimentare ha un impatto determinante sulle pressioni selettive che agiscono sulla specie stessa: nel corso del tempo la capacità cranica di H. erectus varia da circa 750 a oltre 1000 cm3. Se consideriamo che 1000 cm3 è un valore che rientra all’interno della variazione normale degli esseri umani moderni, non desta alcuna sorpresa il fatto che H. erectus sia il primo ominine a sfruttare le sue conoscenze tecnologiche per diffondersi in Eurasia.
La presenza di H. erectus è attestata in Cina 2,1 milioni di anni fa, in Georgia 1,8 milioni di anni fa e in Indonesia più di 1,7 milioni di anni fa. Sarà proprio nel Sud-est asiatico che alcune popolazioni isolate di H. erectus sopravvivranno fino a 50.000 anni fa sviluppando nanismo insulare (H. luzonensis e H. floresiensis).
Sapiens, Neanderthal e Denisova: una famiglia allargata
Mentre H. erectus colonizza l’Eurasia, altri omìnini africani sperimentano altri modi di essere umani. Da un lato abbiamo alcuni rappresentanti dei rami precedenti e caratterizzati dal possesso di caratteri conservati, come H. naledi, una specie che sembra sparire senza lasciare discendenti. Dall’altro abbiamo una serie di specie un tempo raggruppate sotto l’etichetta onnicomprensiva di H. sapiens “arcaici” e che hanno convissuto a lungo in una rete panafricana di relazioni e ibridazioni prima di prodigarsi in ondate di colonizzazione verso gli altri continenti. Si tratta principalmente di H. antecessor e H. heidelbergensis, vissuti tra 700.000 e 300.000 anni fa, tra i quali dobbiamo annoverare l’ultimo antenato diretto e condiviso tra i cosiddetti Denisova (una popolazione ominine che prende il nome da una località dei monti Altai in Asia centrale), H. neanderthalensis (dotato della maggiore capacità cranica media attestata per gli omìnini, con 1450 cm3), e H. sapiens moderno in senso stretto (con una capacità cranica media di circa 1360 cm3). Come H. erectus, anche in questo caso abbiamo esseri umani profondamente legati alla produzione di cultura materiale e al perfezionamento di tecnologie avanzate. Tra queste ultime, migliori lance e strumenti per scavare e rivoluzionarie forme di imbracatura per neonati tramite pelli legate al corpo delle madri conducono a innovative divisioni paritarie del lavoro e a diete più varie.

Episodi di ibridazione tra le varie popolazioni arcaiche di Homo. TRADUZIONE: freccia grigia in alto a destra = episodi di ibridazioni tra popolazioni diverse; Ancestral humans = omìnini umani ancestrali; Estimated number of years ago in alto a destra = Quadro cronologico di riferimento; 2 million = 2 milioni di anni fa; Archaic Homo (“Ghosts”) = popolazioni “fantasma” di Homo “arcaici”; Denisovans = Denisova; Neanderthals = Neanderthal; Modern Humans = esseri umani moderni; Europeans = popolazioni europee; East Asians = popolazioni dell’Asia orientale; Melanesians = popolazioni melanesiane; Africans = popolazioni africane; Present day = oggi. FONTE: Pikaia. Il portale dell’evoluzione, da Science.
Nel corso dei loro incontri e scontri in giro per l’Eurasia, tutti questi gruppi di esseri umani danno vita a ibridazioni tra diverse popolazioni (si tratta difatti di specie geologicamente recentissime la cui somiglianza genetica è valutata essere oltre il 99,8%). Minime percentuali di DNA appartenenti a Denisova e Neanderthal sopravvivono attualmente nelle popolazioni umane viventi.
Nel corso di questi processi di ibridazione, complice una serie di oscillazioni glaciali favorevoli che espongono territori in seguito sommersi dal mare, H. sapiens raggiunge l’Asia oltre 65.000 anni fa, arriva nel Sahul oltre 60.000 anni fa (il paleocontinente che univa l’isola della Nuova Guinea, l’Australia e la Tasmania), si diffonde in Europa tra 40.000 e 20.000 anni fa, e infine giunge nelle Americhe dapprima valicando tra i 20.000 e i 15.000 anni fa un ponte di terra asiamericano ora scomparso detto Beringia e poi attraversando l’Oceano Pacifico per raggiungere l’America meridionale dall’Australasia.
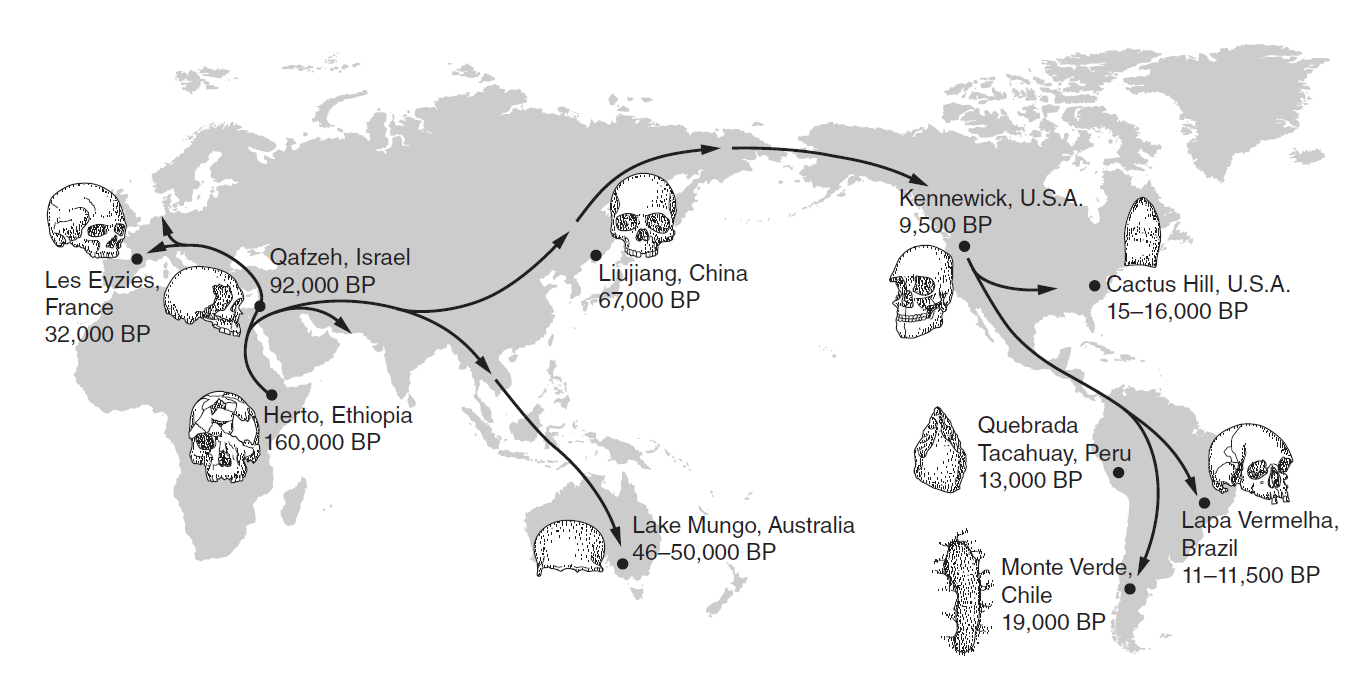
Colonizzazione delle terre emerse da parte di Homo sapiens. Date indicative ricavate da alcuni dei ritrovamenti più significativi. TRADUZIONE: BP (Before Present, ossia prima del 1950 per convenzione cronologica) = anni fa; France=Francia; Israel=Israele; Ethiopia=Etiopia; Brazil=Brasile; Chile=Cile: Peru=Perù; China = Cina. FONTE: Benton, M. (2015). Vertebrate Evolution. 4th edition. Wiley-Blackwell, fig. 11.20, p. 426.
Una moltitudine di antenati
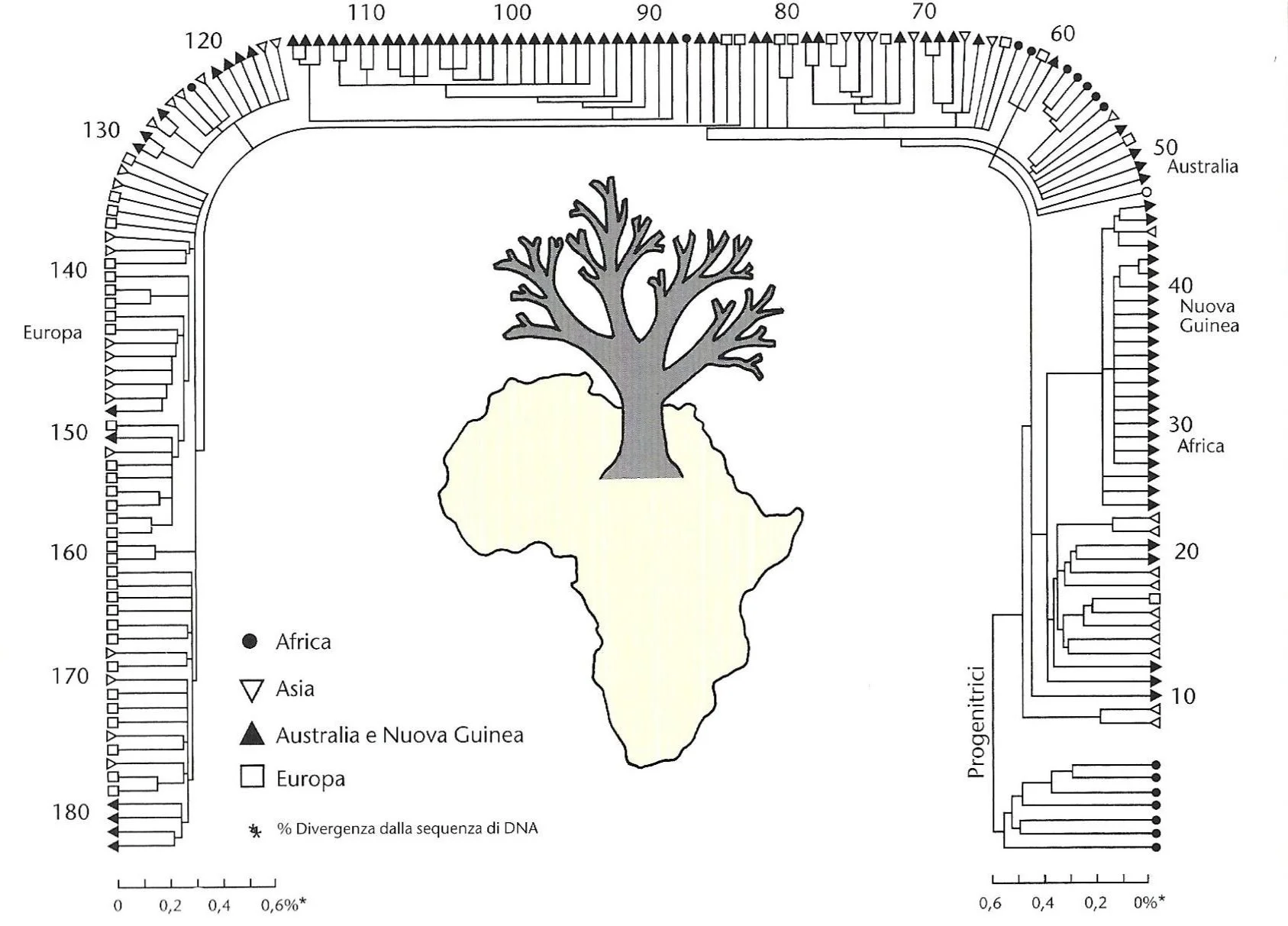
Uno dei primi alberi filogenetici elaborati a partire dall’analisi del DNA mitocondriale. La divergenza genetica indica chiaramente una discendenza a partire da antenati africani (in basso a destra). FONTE: C. Stringer e P. Andrews (2006). Storia completa dell’evoluzione umana. Modena: Logos, p. 179.
Tutte queste specie umane hanno viaggiato portando con sé il proprio patrimonio genetico. Molti di questi viaggiatori hanno avuto figli con gli esseri umani del sesso opposto – anche appartenenti a gruppi diversi – che incontravano nei luoghi che raggiungevano. Tutti però erano membri di popolazioni umane che avevano lasciato l’Africa nel corso di una serie di ondate di esplorazione e colonizzazione.
Ciascuna popolazione delle ultime tre specie di Homo (H. sapiens, H. neandethalensis e i Denisova) era infatti la figlia di lontane diaspore africane. Sappiamo dalle analisi effettuate sul DNA mitocondriale che la linea femminile di H. sapiens risale a circa 200.000 anni fa e ha le sue radici nella zona dell’attuale Botswana, mentre lo studio del cromosoma Y maschile sembra puntare a una linea risalente a circa 400.000 anni fa e localizzata nell’Africa occidentale.

Gli studi paleogenetici rivelano la diversità degli antichi abitanti d’Europa. Il genoma ricavato da una gomma da masticare prodotta dalla corteccia di betulla nella Danimarca di 5.700 anni fa ha permesso di identificare sia il consumatore, una ragazza appartenente a una popolazione di cacciatori-raccoglitori eurasiatici dalla pelle scura, dai capelli castano scuro e dagli occhi chiari, sia i suoi ultimi pasti (nocciole e anatre). I ricercatori sono riusciti anche a stabilire l’intolleranza al lattosio della ragazza. FONTE: © Tom Björklund; illustrazione riprodotta su The Guardian, 17 dicembre 2019; studio originale pubblicato su Nature.
Il quadro che risulta è quello di un continuo rimescolamento di geni e di culture, tant’è che nessun gruppo può rivendicare una continuità ancestrale con un certo luogo. Nonostante certe pretese campanilistiche nate durante i processi politici di invenzione delle tradizioni nazionali tra l’Ottocento e il Novecento del secolo scorso, la paleogenetica sta mostrando come i profili genetici delle popolazioni umane che vivono oggi in una data zona spesso non corrispondano a quelli degli esseri umani che occuparono nel passato profondo quella stessa area.
Inoltre, nessuno può dire di essere un essere umano “puro” dal momento che nelle varie regioni della Terra il DNA degli esseri umani moderni conserva le tracce dei gruppi umani distinti fra loro che si sono più volte incrociati in epoche diverse. La paleogenetica sta inoltre rivelando l’esistenza di “popolazioni fantasma” di H. sapiens “arcaici” per ora note esclusivamente attraverso le porzioni di DNA sopravvissute negli esseri umani attuali.
L’espressione parafrasa una celebre frase del poeta statunitense Walt Whitman (1819-1892): “Mi contraddico? Certo che mi contraddico: sono vasto, contengo moltitudini”.
Tuttavia, anche se queste popolazioni per ora ignote sono scomparse, così come i Denisova e i Neanderthal (l’estinzione di questi ultimi risale a soli 27.000 anni fa), alcuni dei loro geni sopravvivono negli esseri umani moderni. Possiamo ben dire di contenere “moltitudini”* dentro di noi.
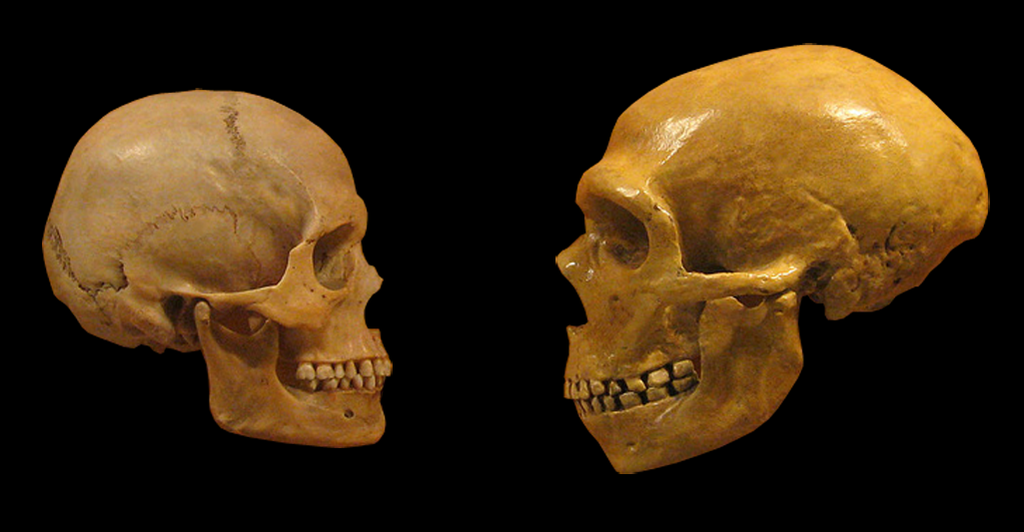
Confronto tra un cranio di Homo sapiens moderno (A SINISTRA) e H. neanderthalensis (A DESTRA). FONTE: hairymuseummatt (original photo), DrMikeBaxter (derivative work) [CC 2.0], da Wikipedia.